
Induced Pluripotent Stem Cells (iPSCs) have a long and rich development history, where the ideas leading up to Yamanaka’s work date back to before the 1960s. In this edition of the REPROCELL blog we discuss some of those discoveries. We also mention some recent developments with an emphasis on reprogramming.
1. History
1.1. The Origins of Cellular Reprogramming (1962)
Well before the development of iPSC lines came John Gurdon’s 1962 Nobel prize winning experiments on the genesis of Xenopus Laevis frog model. These experiments were published in the Journal of Embryology and Experimental Morphology (known today as Development).1 They involved using the nuclei of terminally differentiated intestinal epithelial cells and transferring them into enucleated eggs – that is, somatic cell nuclear transfer. In this work, Gurdon both developed the technique used in reproductive cloning – the one used to create Dolly the sheep, and, also demonstrated that the genetic material contained in a cell is preserved over the differentiation process.2
1.2. Challenging Developmental Dogma
Reflecting on these experiments nearly 50 years later, Gurdon noted that the experiments highlighted how as cells differentiate their nuclei are less capable of forming developed embryos. This feature, he says, was reminiscent of reprogramming resistance seen in some iPSC experiments today. His work was groundbreaking in that the understanding of how phenotypic diversity emerged in a complete organism failed to account for whether irreversible mutational events were required. What Gurdon showed was the possibility of returning somatic cells to their pluripotent state.1 This challenged August Weismann’s Germ Plasm Theory (1892) and Conrad Waddington’s epigenetic landscape (1957) both theories that posited somatic cells were restricted to their phenotype by irreversible modifications to genetic material.3,4 Knowing this, it makes sense to ask what exactly is the molecular machinery involved in the differentiation? And, in what ways can it be manipulated? These questions, however, would wait until the existence of suitable culture techniques (Figure 1).
1.3. The First Embryonic Stem Cell Cultures (1981)
The next major milestone came 19 years later, when Martin Evans and Matthew Kaufman reported the first successful culture of pluripotent mouse embryonic stem cells. These cells were harvested from in vitro cultures of mouse blastocytes; until then, scientists were unable to grow stem cells in culture. Instead, pluripotent lines needed to be harvested after teratocarcinoma formation in vivo.
Actually, teratocarcinoma cell lines and their embryonal carcinoma stem cells were thought to resemble human embryonic cells and serve as an in vitro substitute.6 In their 1981 paper, Evans and Kaufman noted three factors they considered to be critical for the success of culture – those were: the exact stage at which pluripotent cells capable of culture are found in the embryo, the explantation of a sufficient quantity of these cells, and culture conditions that promote multiplication over differentiation. Their paper reports that blastocysts were cultured in groups of roughly 6 embryos in small drops of medium under paraffin oil for 4 days.
The authors noted that inner cell mass cells turned into egg cylinder-like structures (i.e., a group of round cells surrounded by the endodermal cells). These egg cylinder structures were passaged onto gelatin precoated Petri dishes with mitomycin C-inactivated STO fibroblasts using trypsin. Remarkable to their newly established cell line was a normal karyotype (with Figure 2 of the paper being a normal XY karyotype), something they reported that no other pluripotent line in their lab had achieved.5
1.4. The First Human Embryonic Stem Cells (1998)
The first human embryonic stem cell lines, however, were developed by Thomson et. al. 17 years later, who in their abstract reported that their cells could be cultured for 4 to 5 months and still form both trophoblast and derivatives of the three embryonic germ layers.7 This achievement marked a turning point for regenerative medicine by providing a renewable source of human pluripotent cells for research.
1.5. Ethical and Political Debate
Shortly after, controversy began. A 1998 Science Adviser newsletter by Eliot Marshall discussed the impact of Thomson’s work. Marshall reports that the work was thought of as a substantial breakthrough by scientists in the field. However, Marshall mentions that Congress passed a bill placing restrictions on the use of U.S. funds on research relating to the use of embryos. Marshall described that the bill forced Thomson to set up a separate research space to block his work from any public funding sources. According to the newsletter, Thomson was funded mostly by a company in exchange for the license to his discovery. The letter concludes with a remark on the worry tied to the discovery’s nebulous legal status.8
1.6. U.S. Stem Cell Policy and NIH Guidelines
In 2009, Executive Order 13505 was issued by President Barack H. Obama legalizing stem cell (e.g., hESC – human Embryonic Stem Cell) research in the U.S. after 49,000 comments from various people and groups.9 Published in the Federal Register (74 Fed. Reg. 18578 (April 23, 2009)) were guidelines for implementing the executive order at the NIH. Of note is how the hESCs used for research need to be from embryos derived from in vitro fertilization programs – from embryos formerly intended for use in human reproduction. The comment also included a list of assurances that need to be met, like, that the donor’s decision to create the embryo and the subsequent decision to donate the embryo for research purposes are distinct events, or, that the reproductive clinical care physician and researcher are not the same person. The comment defined embryonic stem cells as cells from the human embryo, capable of dividing for prolonged periods in culture without differentiation, and have a known capacity to turn into cell/tissue of the three primary germ layers. The Federal Register entry makes clear that the stem cells in question are not themselves human embryos.10
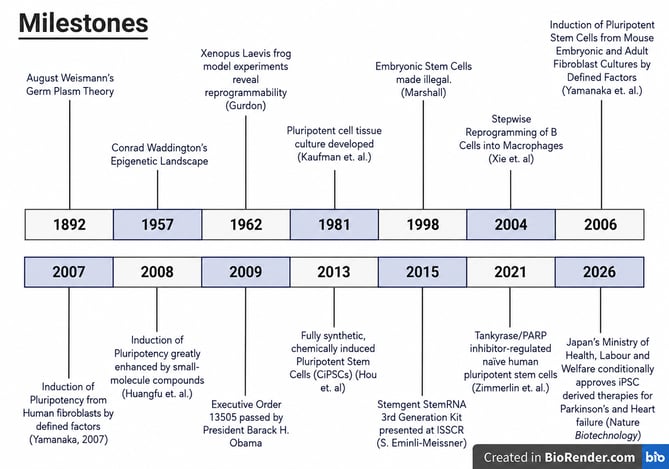
Figure 1. Timeline highlighting major scientific and clinical milestones that shaped the development of induced pluripotent stem cell (iPSC) technology.
2. Transcription Factors Determine Lineage
2.1. Transcription Factors Control Cell Fate
The next set of key events involve experiments which elucidate the role of transcription factors (TFs) in cell fate. TFs are proteins which regulate the transcription of DNA into RNA. What we know today is that by expressing them strategically, cell fate can be controlled.
One study central to the development of this understanding retrovirally expressed these transcription factors in mature B cells, then seeded them on stroma. After 3 days, 31% of cells became Mac-1+, as indicated by flow cytometry.11 Mac-1, otherwise known as macrophage antigen 1 is an integrin responsible for recognizing and binding to molecules on the surface of invading bacteria. Mac-1 is found on T lymphocytes, B cells, and natural killer cells – particularly, lymphoid cells.12 Myeloid cells, on the other hand, include macrophages, granulocytes, megakaryocytes, and erythrocytes13 – components of the innate immune system.
2.2. Reprogramming B Cells into Macrophages
What the paper demonstrated was C/EBPα/β inhibits Pax5, a B-cell commitment factor, causing CD19 downregulation, and the authors also showed that Mac-1 upregulation happens through a synergy with endogenous PU.1 and C/EBPα, where, this phenomena and the CD19 downregulation process occur in an uncoupled manner.11
It was generally known that a tightly regulated transcription factor network coordinated the unique gene expression patterns in cells of the hematopoietic lineages, but, that the TFs responsible for the lymphoid/myeloid cell differentiation fate fork were unknown.11 This is important because understanding this fork meant understanding the machinery involved in the B-cell/macrophage differentiation process.
Prior work involving use of the raf/ras oncogenes, and a study involving a form of the M-CSF receptor for reprogramming revealed that the resulting cells featured immunoglobulin rearrangements which were the same as the original B lineage cells.11 The authors argued that this implies reprogramming features a complete halt of the B cell specific gene expression program, and subsequent implementation of a macrophage-specific program.11
2.3. Experimental Design and Validation
For their experiment, relevant cells were transduced with retroviruses where cDNAs of PU.1, C/EBPα, and Pax5 were cloned by PCR into a pMIG vector.11 Full length C/EBPβ, however, was gifted by another scientist and inserted to the same vector at a different site.9
The B cell progenitors used in this study were obtained from the marrow of C57BI/6J mice and purified with magnetic beads for those which were CD19+. Flow cytometry indicated that the cells infected with control virus remain Mac-1 negative but retain CD19 expression (denoted Mac-1-/CD19+), whereas, the cell population infected with the vector containing C/EBPα were 60% Mac-1+/CD19- (from Figures 1C and 1D).11
To rule out contamination by macrophage in the initial culture, the authors developed a so-called “Cre Lox Lineage Ancestry approach.” In this assay, mice were bred so that EYFP could be observed when Cre was active. According to Figures 2B and 2C, cells gated EYFP- were Mac-1+ while EYFP+ cells were CD19 positive but not Mac-1+, indicating that their system successfully screens for the initial B-cells. For cells infected with virus containing C/EBPα and hCD4 Mac-1 was activated and CD19 was downregulated in 65% of hCD4+ EYFP+ cells, meaning that their initial results on reprogramming were not due to contaminating macrophages.11
2.4. Mechanistic Insights
For the interested reader, the result section of the paper features 13 sections, where gene profiling is considered, as is, whether endogenous PU.1 is required in Mac-1 activation/CD19 downregulation – this was shown by infecting a PU.1 deficient pre-B-Cell line with C/EBPα, or PU.1, or both and assessing. They found that CD19 downregulation happens to the same extent whether PU.1 was present or not, but that Mac-1 activation requires PU.1.11
What this work represents is a step beyond somatic cell nuclear cloning with respect to the ability to ‘reprogram’ cells – further dismantling the assumption that cell differentiation is irreversible. It provides a conceptual background for the work of Yamanaka and Takahashi, but in the context of immune differentiation.
3. Discovery of Induced Pluripotent Stem Cells
3.1. Searching for the Factors of Pluripotency
Yamanaka and Takahashi’s work began with 24 transcription factors known to be involved with embryonic stem cell (ES cell) pluripotency. After cloning the factors into retroviral vectors, they used mouse embryonic fibroblasts to combinatorially screen them for the key four (Oct4, Sox2, Klf4, and Myc).14 These became known as the Yamanaka factors and enabled the lab to later develop iPSCs from human fibroblasts – the iPSC technology.15
Their paper began with a statement on how the transcription factors responsible for pluripotency induction were unknown, despite the fact that scientists knew how to achieve an embryonic cell like state by applying somatic cell nuclear transfer. They asserted that though Human ES cells have the potential to treat diseases such as Parkinson’s and spinal cord injuries, ethical issues on the use of human embryos and stem cell transplant rejection make generating autologous pluripotent lines desirable.14
Their paper referenced current literature on what TFs were known and relevant to their experiments (among which were Oct3/4, Sox2, c-myc, Klf4, and b-catenin, etc.), citing 14 studies in this section.14 Yamanaka and Takahashi also included a dominant-negative mutant of Grb2∆SH2 – i.e., the Growth factor receptor-bound protein 2 (Grb2) with a certain SH2 domain.14,16
3.2. Screening the Original 24 Factors
Their experiment involved screening for pluripotency by applying G418 resistance.14,17 They achieved G418 resistance by knocking in a fusion of the β-galactosidase and neomycin resistance genes into the mice’s Fbx15 locus, a gene needed for pluripotency maintenance and mouse development.14,18 While they didn’t attain drug resistance with any one TF, transducing all 24 candidates at the same time produced 22 drug resistant colonies. Repeating the experiment and using RT-PCR indicated that the cells resembled ES cells in doubling time and gene expression, where, they obtained 29 drug resistant colonies the second time and selected four clones for RT-PCR and doubling time. They designated these cells “pluripotent stem cells induced from MEFs by 24 factors” – or, iPS-MEF24 (Figure 2).14
 Figure 2. iPSC reprogramming methods and pluripotency assessment. Overview of integrating and non-integrating reprogramming strategies and the assays commonly used to confirm iPSC pluripotency. Reproduced from Aboul-Soud et al. 12 under CC-BY 4.0.
Figure 2. iPSC reprogramming methods and pluripotency assessment. Overview of integrating and non-integrating reprogramming strategies and the assays commonly used to confirm iPSC pluripotency. Reproduced from Aboul-Soud et al. 12 under CC-BY 4.0.
3.3. Identification of the Four Yamanaka Factors
After, they omitted the TFs, one at a time, from the transduction pool.What this revealed was 10 individual factors which produced no colonies when absent – further, transduction with these 10 simultaneously produced more ES-cell like colonies than all 24 genes.14
They then screened again by omitting one a time from the 10 critical transcription factors. This time, Oct3/4, Klf4, Sox2, and c-Myc were the four deemed necessary. Using two transcription factors alone failed to produce drug resistant colonies, but certain combinations of three factors produced drug resistant cells (Oct3/4, Klf4, Sox2, and, Oct3/4, Klf4, c-Myc) though, these colonies had morphologies different from the iPSCs generated in the other trials.14
3.4. Characterisation of the Newly Generated iPSCs
RT-PCR and Chromatin immunoprecipitation assays were performed on the iPS-MEF10 and iPS-MEF9 clones, where the authors found that the cells had most of the marker genes and that Oct3/4 and Nanog promoters had upregulated acetylation of histone H3 and downregulation of demethylation of histone H3’s lysine 9.14,19
For the ChIP protocol used in this paper, the authors reference a previous paper involving the roles of Sox15 and Sox2 in transcriptional control.14, 20 The authors assessed the cells in several other experiments, looking at global gene-expression profiles, pluripotency by teratoma formation, embryoid body formation, and how the four key transcription factors fared in generating G418 resistance in tail-tip fibroblasts.14 The reader is encouraged to review their 2006 paper for more detail.
3.5. Limitations of Early Reprogramming
One of the limitations tied to their work, they mention, was low reprogramming efficiency. They propose that the four factors need to be expressed in narrow ranges for iPSC induction to go through.14 That is, it may have been that a small fraction of the cells expressed them in the way needed, resulting in low iPSC derivation.
They also mention that certain spontaneous chromosomal alterations may be required for iPSC induction. These, among other suggestions, preclude that the small number of iPSCs observed were from pluripotent cells residing in the initial mouse skin cell culture expanding (of which, 0.067% should be stem cells).14
3.6. Parallel Discovery of Human iPSCs
The work of Thompson in the United States was conducted in parallel and involved the OCT4, SOX2, NANOG, and LIN28 factors. Both scientists were, independently responsible for the discovery of human induced pluripotent stem cells.21
4. Improved Reprogramming Methods
4.1. Small Molecule Approaches
Over time, reprogramming methods have become more refined – including small- molecule and small-molecule-assisted methods.21 Relying on small molecules relieves some of the strain associated with genetic manipulation methods.22 For example, one group was able to show that small molecule histone deacetylase (HDAC) inhibitors improve reprogramming efficiency over 100-fold.Their work – published to nature biotechnology – used an Oct4-GFP reporter to assess if the small molecules involved in chromatin modifications can influence reprogramming.23 GFP reporter systems involve fusing GFP to a gene’s promotor so that GFP expression is observed when the gene is transcribed.
Other examples of this tool being used in stem cell biology include as a reporter system deriving air way basal stem cells, in modeling serotonergic neuron development, and in tracing oncogene driven remodeling of the intestinal stem cell niche. 24,25,26 They found out that suberoylanilide hydroxamic acid, trichostatin A, and valproic acid (VPA) boosted reprogramming efficiency, where, VPA had the strongest effect.23
4.2. Chemically Induced Pluripotent Stem Cells (CiPSCs)
Chemical reprogramming continued to advance with the development of a seven-small-molecule protocol capable of converting mouse fibroblasts into CiPSCs. A seven-small molecule process was published which allowed the authors to reprogram mouse fibroblasts to CiPSCs.22 The group previously found that valproic acid, CHIR99021, and tranylcypromine could replace all but Oct4 in the reprogramming process.27 They then showed that forskolin, 2-methyl-5-hydroxytryptamine, and D4476 could replace Oct4 in subsequent work, resulting in a fully synthetic 7-compound reprogramming methodology. According to the abstract, their cells were like embryonic stem cells in gene expression profiles among other properties with 0.2% of the cell population demonstrating pluripotency.22
4.3.Expanding Chemical Reprogramming Beyond Fibroblasts
In other work, Ye et. al assessed chemical reprogramming methods for non-fibroblast cell lines. As representatives of the ectoderm and endoderm lineages, they successfully reprogramed neural stem cells and intestinal epithelial cells. In this work, they developed a lineage tracing system using fibroblast specific protein-1. They also used an Oct4-GFP reporter system when reprogramming NSCs – this allowed them to keep track of where in the reprogramming process the NSCs were.28
4.4. Viral Reprogramming Methods
Beyond small molecule reprogramming, the conventional reprogramming methods, (like the one used in the Xie et. al paper, and by Yamanaka) involve viral transduction, where a virus carrying the genetic reprogramming information delivers the material. Viral vectors can be integrating or non-integrating. The main difference is that non-integrating vectors are cleared from cells as they proliferate. Scientists deliver Oct3/4, Sox2, Klf4, and c-Myc (or, OSKM) expression cassettes virally because this method is efficient. The lentivirus and retroviral delivery methods are integrating and result in stable expression of the factors, but are limited by insertional mutagenesis and unwanted transgene reactivation. Adenoviruses, Adeno-associated viruses, and Sendai viruses are non-integrating viruses.21
4.5. mRNA-Based Reprogramming
Alternatively, one can deliver without integration mRNAs of the transcription factors using lipid nanoparticles. The StemRNA™ 3rd generation reprogramming kit, for example, offers this type of protocol. The kit (ReproCELL, cat. no. 00-0076) uses six RNAs for reprogramming, immune-ablating RNAs (using E3, K3, and B18), and a reprogramming-associated microRNA cocktail in a manner which saves 4-8 weeks’ worth of time dedicated to vector removal screening. The kit relies on the Lipofectamine® RNAiMAX™ Transfection Reagent (ThermoFisher Scientific, 13778030), a cationic lipid transfection reagent. In 10-14 days, primary iPSC colonies can be seen depending on the starting cell type. Whereas the work of Yamanaka and Takahashi featured murine embryonic fibroblasts for reprogramming, methods are now generalizable to other cell types. Certainly, urine derived progenitors and endothelial progenitors derived from blood can be reprogrammed.29
5. Conclusion
Nearly 20 years has passed since their discovery, induced pluripotent stem cells will continue to play a role in regenerative medicine, oncology, and disease modelling. Not only will our understanding of pluripotency and the underlying biology expand but our capacity to leverage pluripotent lines therapeutically will too. Consider, for example the discovery of Tankyrase/PARP inhibitor-regulated naïve human pluripotent stem cells (TIRN-hPSC), a new variety of human stem cell capable of differentiating into embryonic-like pericytic-endothelial “naïve” vascular progenitors (N-VP).30 Reversion of hPSCs to this state fixes dysfunctional epigenetic donor cell memory – i.e., residual epigenetic machinery due to differentiation from donor cell, lineage-primed gene expression, and possibly disease-associated aberrations in differentiated progeny.30 Further, this line of PSC has already shown use in revascularizing the ischemic retina, starting from diseased (e.g., diabetic) donor material.31 This exciting discovery offers hope for those suffering from diabetic retinopathy. Clinical trials involving iPSCs are already underway and will continue to progress, barring any safety concerns.
Readers are encouraged to continue active lines of inquiry and REPROCELL inc., will be available with the products and services you need to make the most out of your projects.
References
- Gurdon, J. B. The cloning of a frog. Development, 140 (12): 2446–2448. (2013).
- Gurdon, J. B. The Developmental Capacity of Nuclei taken from Intestinal Epithelium Cells of Feeding Tadpoles. Development 1; 10 (4): 622–640. (1962).
- S. AE. Das Keimplasma Eine Theorie der Vererbung. Nature, 47(1212):265-266.(1893).
- Waddington, C. H. The Strategy of the Genes. 0 ed. Routledge, (2014).
- Evans, M. J. and Kaufman, M. H. Establishment in Culture of Pluripotential Cells from Mouse Embryos. Nature, 292 (1981).
- Andrews, P. W. Human Teratocarcinomas. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer, 948 (1): 17-36. (1988).
- Thomson, J. A., Sander, S. S., Michelle, A. W., Jennifer, J. S. Vivienne, S. M. and Jeffrey, M. J. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science, 282, no. 5391: 1145–47.(1998).
- E. Marshall. https://www.science.org/content/author/eliot-marshall
- Kington, R. S. NIH Guidelines for Human Stem Cell Research. National Institutes of Health: Turning Discovery into Health. (2009).
- National Institutes of Health. Draft National Institutes of Health Guidelines for Human Stem Cell Research Notice. Federal Register, The daily journal of the United States Government. (2009).
- Xie, H., Min, Y., Ru, F. and Thomas, G.. Stepwise Reprogramming of B Cells into Macrophages. Cell, 117, no. 5: 663–76. (2004).
- Springer, T., Galfré, G., Secher, D.S. and Milstein, C. Mac-1: a macrophage differentiation antigen identified by monoclonal antibody. Eur J Immunol. 9(4):301–306. (1979).
- MacDonald, R. Immunology: What are myeloid cells and how are they identified?. Cell Signaling Technology. (2025).
- Takahashi, K. and Yamanaka, S. Induction of Pluripotent Steb m Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell, 126, no. 4: 663–76. (2006).
- Cho, H.E., Lee, S., Seo, J. H., Kang, S.W., Choi, W.A. and Cho, S.R. In Vivo Reprogramming Using Yamanaka Factors in the CNS: A Scoping Review. Cells, 13(4):343. (2024).
- Wang, D., Liu, G., Meng, Y., Chen, H., Ye, Z. and Jing, J. The Configuration of GRB2 in Protein Interaction and Signal Transduction. Biomolecules, 14(3):259. (2024).
- InvivoGen. G418 (Geneticin). Available from: https://www.invivogen.com/g418. Accessed July 16, 2026.
- Tokuzawa, Y., Kaiho, E., Maruyama, M., Takahashi, K., Mitsui, K., Maeda, M., Niwa, H. and Yamanaka, S. Fbx15 is a novel target of Oct3/4 but is dispensable for embryonic stem cell self-renewal and mouse development. Mol Cell Biol, 23(8):2699-2708. (2003).
- Cell Signaling Technology. Overview of Chromatin Immunoprecipitation (ChIP). Accessed July 16, 2026.
- Maruyama, M., Ichisaka, T., Nakagawa, M. and Yamanaka, S. Differential roles for Sox15 and Sox2 in transcriptional control in mouse embryonic stem cells. J Biol Chem. 280(26):24371-24379. (2005).
- Cerneckis, J., Cai, H. and Shi, Y. Induced pluripotent stem cells (iPSCs): molecular mechanisms of induction and applications. Signal Transduction and Targeted Therapy, 9(1):112. (2024).
- Hou, P., Li, Y., Zhang, X., Liu, C., Guan, J., Li, H., Zhao, T., Ye, J., Yang, W. and Deng, H. Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds. Science, 341(6146):651-654.
- Huangfu, D., Maehr, R., Guo, W., Eijekelenboom, A., Snitow, M., Chen, A.E. and Melton, D.A. Induction of pluripotent stem cells by defined factors is greatly improved by small-molecule compounds. Nat Biotechnoly, 26(7):795-797. (2008).
- Hawkins, F.J., Suzuki, S., Beermann M.L., Barillà, C., Wang, R., Villacorta-Martin, C., Berical, A., Jean, J.C., Le Suer, J., Matter, T., SImone-Roach, C., Tang, Y., Schlaeger, T.M., Crane, A.M., Matthias, N., Huang, S.X.L., Randell, S.H., Wu, J., Spence, J.R., Carraro, G., Stripp, B.R., Rab, A., Sorsher, E.J., Horani, A., Brody, S.L., Davis, B.R. and Kotton, D.M. Derivation of Airway Basal Stem Cells from Human Pluripotent Stem Cells. Cell Stem Cell. 28(1):79-95.e8. (2021).
- Pacini, G., Marino, A., Migliarini, S., Brilli, E., Pelosi, B., Maddaloni, G., Pratelli, M., Pellegrino, M., Ferrari, A. and Pasqualetti, M. A Tph2GFP Reporter Stem Cell Line To Model in Vitro and in Vivo Serotonergic Neuron Development and Function. ACS Chem Neurosci, 8(5):1043-1052. (2017).
- Yum, M.K., Han, S., Fink, J. and Wu, S.S., Dabrowska, C., Trendafilova, T., Mustata, R., Chatzeli, L., Azzarelli, R., Pshenichnaya, I., Lee, E., England, F., Kim, J.K., Stange, D.E., Philpott, A., Lee, J., Koo, B. and Simons, B.D. Tracing oncogene-driven remodelling of the intestinal stem cell niche. Nature, 594(7863):442-447. (2021).
- Li, Y., Zhang, Q., Yin, X., Yang, W., Du, Y., Hou, P., Ge, J., Liu, C., Zhang, W., Zhang, X., Wu, Y., Li, H., Liu, K., Wu, C., Song, Z., Zhao, Y., Shi, Y. and Deng, H. Generation of iPSCs from mouse fibroblasts with a single gene, Oct4, and small molecules. Cell Res, 21(1):196-204. (2011).
- Ye, J., Ge, J., Zhang, X., Cheng, L., Zhang, Z., He, S., Wang, Y., Lin, Y., Yang, W., Liu, J., Zhao, Y. and Deng, H. Pluripotent stem cells induced from mouse neural stem cells and small intestinal epithelial cells by small molecule compounds. Cell Res, 26(1):34-45. (2016).
- REPROCELL. StemRNA™ 3rd Gen Reprogramming Kit [brochure]. (2023).
- Zimmerlin, L., Park, T.S., Bhutto, I., Lutty, G. and Zambidis, E.T. Generation of Pericytic-Vascular Progenitors from Tankyrase/PARP-Inhibitor-Regulated Naïve (TIRN) Human Pluripotent Stem Cells. Methods Mol Biol, 2416:133-156. (2022).
- Park, T.S., Zimmerlin, L., Evans-Moses, R., Thomas, J., Huo, J.S., Kanherkar, R., He, A., Ruzgar, N., Grebe, R., Bhutto, I., Barbato, M., Koldobskiy, M.A., Lutty, G. and Zambidis, E.T. Vascular progenitors generated from tankyrase inhibitor-regulated naïve diabetic human iPSC potentiate efficient revascularization of ischemic retina. Nat Communications, 11 (1):1195. (2020).

About the author
Yousefsaid Abderrahim, Research Associate, REPROCELL USA
Yousefsaid Abderrahim studied Bioengineering at the University of Maryland, College Park with an emphasis on Biotechnology and Therapeutics Engineering.
Subjects we write about
- 3D Cell Culture
- Cell Culture
- Central Lab Services
- Clinical Capabilities
- Disease Modeling
- Drug Discovery
- Gene Editing
- Genomic Services
- GMP
- Human Tissue Samples
- Human Tissue Testing
- IBD
- Life Sciences
- Master Cell Banks
- Neurons
- Oligonucleotide Synthesis
- Pharmacology-AI
- Precision Medicine
- Product Catalog
- Regenerative Medicine
- Respiratory Disease
- Safety Pharmacology
- Skin Disease
- Stem Cells

.png?width=400&height=225&name=hubspot_featured_image_1200x628%20(1).png)



